
by Sara Murphy and Jiayi Xu
Introduction
Moraxella catarrhalis is a Gram-negative bacterium that was first discovered in 1986. It is found only in humans and until recently, it was considered to be a harmless colonizer of the nose, mouth and throat. However, M. catarrhalis is now recognized as an important cause of ear infections in children under the age of two, and lower respiratory tract infections in adults with compromised immune systems.
Disease
M. catarrhalis is primarily transmitted through mucosal secretions between family members, children in daycares, and between hospital patients in the winter. When the bacterium comes in contact with an infant, it attaches itself to the upper respiratory airways, specifically the nasopharynx. Once M. catarrhalis has invaded small airway epithelial cells, it moves from the nose or mouth to the middle ear by means of the Eustachian tube and causes inflammation along the way (Figure 1). The resulting congestion of fluid due to the narrowed Eustachian tube causes pressure to build in the middle ear, and ultimately results in pain. This is referred to as otitis media.
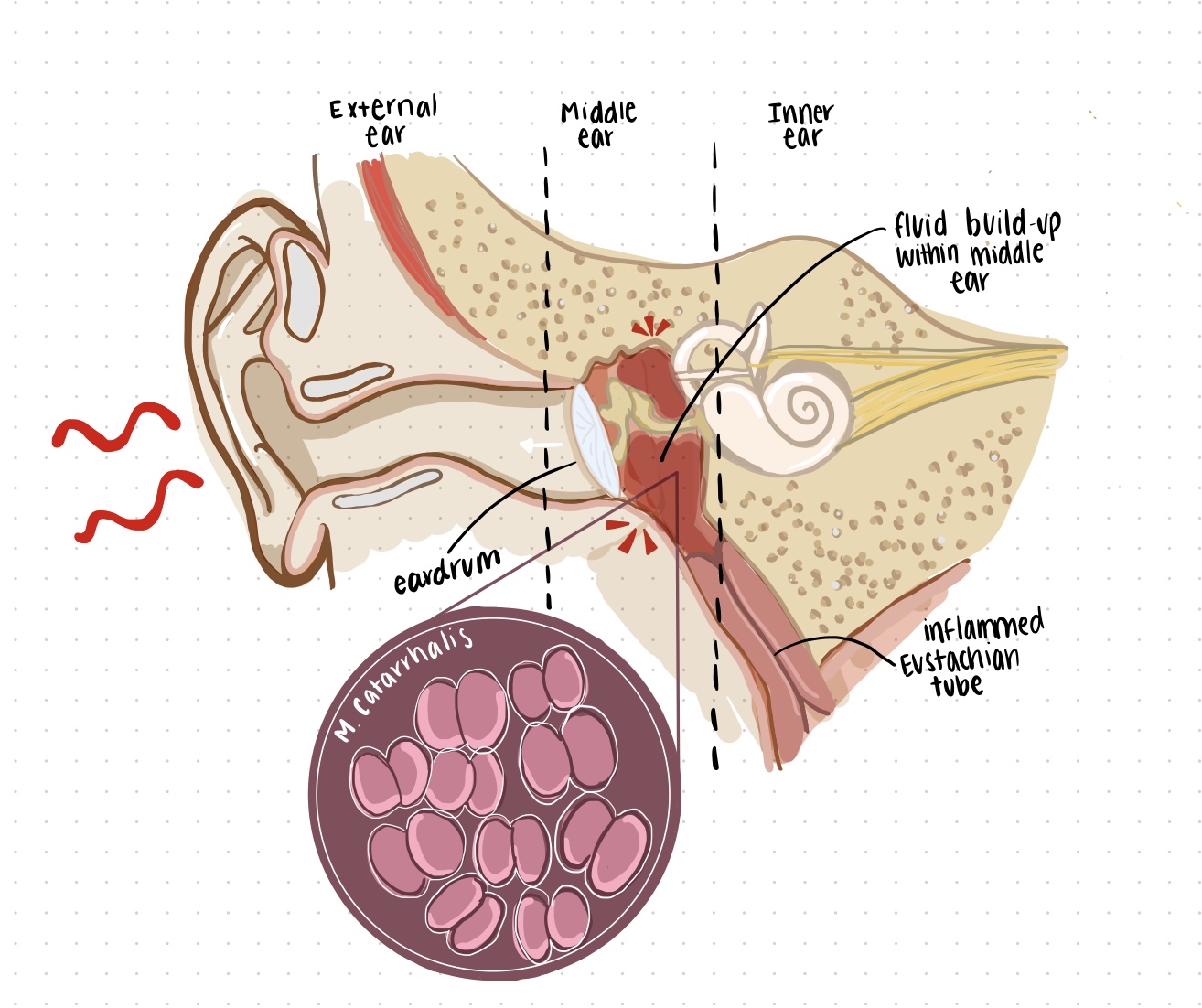
In the case of immunocompromised individuals or those living with chronic disease, M. catarrhalis infection has been shown to increase the severity of existing chronic obstructive pulmonary disease (COPD). When M. catarrhalis invades specific cells within the lungs (i.e. bronchial and alveolar epithelial cells), it contributes to increased inflammation and aggravates the disease symptoms. This includes reduced airflow during respiration, a worsening cough and pulmonary emphysema. However, very little is actually known about how M. catarrhalis leads to exacerbations in patients with COPD.
Epidemiology
Moraxella catarrhalis causes approximately 15-20% of the total acute otitis media cases in infants with up to 80% of children having been colonized before their second birthday. However, the rate of colonization of the nasopharynx decreases considerably in adulthood. Nevertheless, there are roughly 2-4 million worsening cases each year of chronic obstructive pulmonary disease (COPD) associated with M. catarrhalis infection in the United States. Despite this extreme number, the actual rate of disease is likely higher as M. catarrhalis can sometimes be mistaken for Neisseria species in cultures taken from the lungs.
In most cases, M. catarrhalis lives as a natural inhabitant of the human respiratory tract without causing problems. However, under certain circumstances it can become pathogenic. This is highly dependent on age and the development of the immune system, as colonization rates lower significantly as individuals get older. Within similar age groups, the differences in colonization rates can be attributed to genetic predisposition, the number of household members, personal hygiene, and other environmental factors such as living with smokers.
Virulence Factor
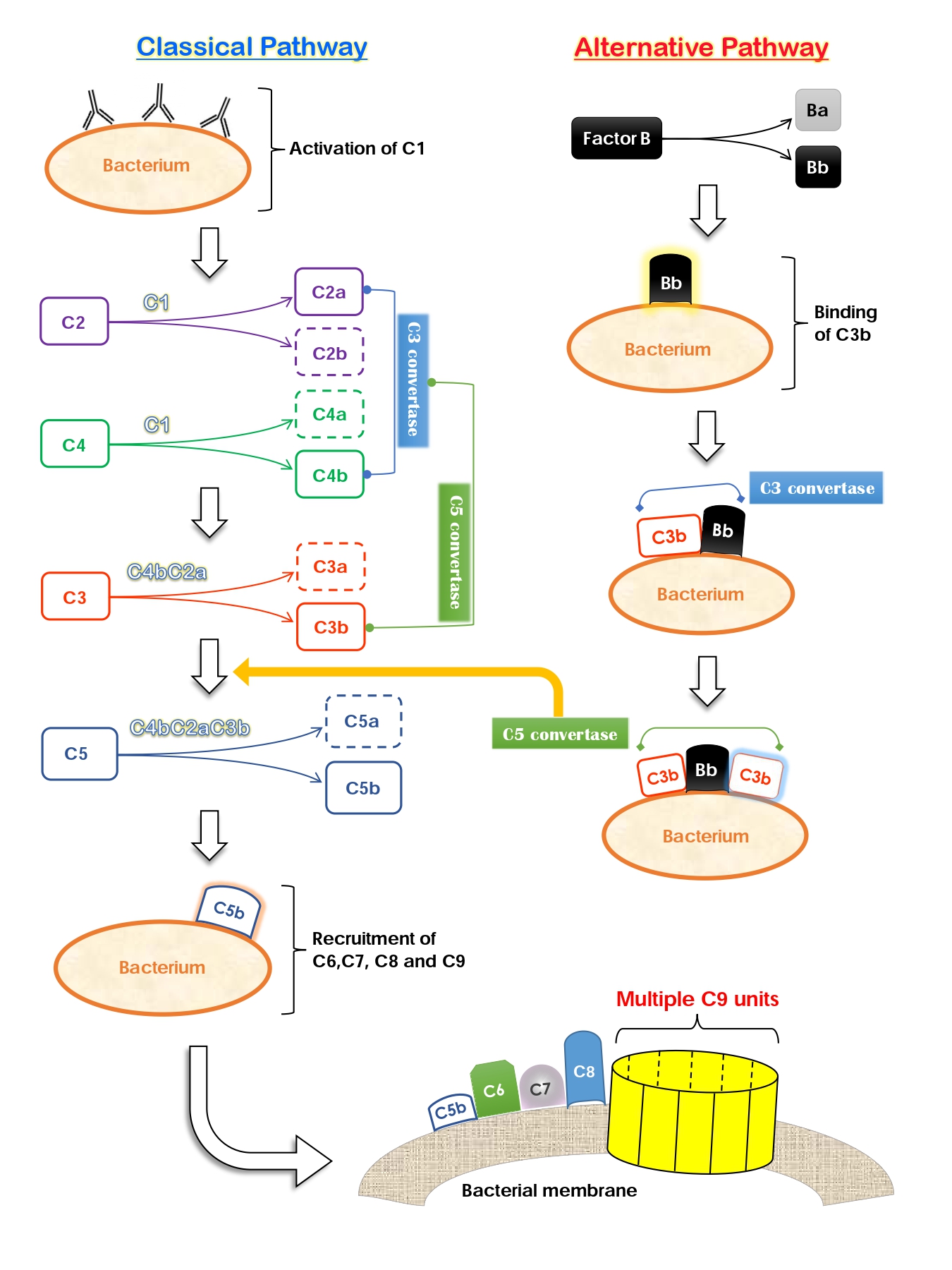
Virulence factors are tools used by a pathogen to enhance its ability to colonize and cause damage to its host. One of the virulence factors found in M. catarrhalis is the Ubiquitous Surface Protein (Usp) family. These proteins are located in the outer membrane of the bacteria. Usp had been shown to confer serum resistance, where the pathogens evade bactericidal activities of the human serum, particularly that mediated by the Complement System. The system includes a series of soluble and membrane-bound proteins, each playing a different role. For example, C3a is pro-inflammatory; whilst C3b strengthens phagocytic activities. While C5a attracts immune cells to the site of infection, C5b is responsible for recruiting the membrane attack complex (MAC), which consists of C6, C7, C8 and multiple C9 subunits. As its name suggests, the MAC lyses bacterial cells and thus contributes directly to the bactericidal function of the Complement System.
The Complement System can be activated in three ways, two of which are the Classical and the Alternative pathway (Figure 2). The classical pathway is initiated by antibodies and results in the production of C2a and C4b. The two then form the C3 convertase and yields C3a and C3b, which also joins the C4bC2a complex to give rise to the C5 convertase. One of the enzyme’s products, C5b, coordinates assembly and binding of the MAC to the bacterial membrane. Unfortunately, M. catarrhalis is equipped with virulence factors that can interfere with this pathway. Two types of the Usp family, UspA1 and UspA2, are able to bind to the C4b-binding protein (C4BP). C4BP is a regulator of the classical complement activation pathway; it promotes degradation of the C3 convertase. Therefore, M. catarrhalis achieves serum resistance by “recruiting” inhibitory host proteins to reduce the amount of downstream proteins necessary for eventual activation of the MAC.
To make things worse, this pathogen species is also able to interfere with the alternative complement activation pathway. Normally, bacterial surface structures will be recognized and bound by the host serum protein Bb. Bb will attract C3b and the two form a C3 convertase, which then amplifies C3b production as well as facilitates subsequent reactions (for MAC activation). However, UspA1 and UspA2 are able to bind to C3 which prevents it from producing C3a and C3b. Thus, binding of the two proteins decreases the quantity of precursor molecules required for the alternative method of complement activation. Hence, UspA1 and UspA2 are two critical virulence factors in M. catarrhalis that protect the bacteria from being lysed by their host’s serum proteins.
Treatment
Almost all strains of M. catarrhalis are resistant to β-lactam antibiotics such as penicillin, amoxicillin and ampicillin. These antibiotics contain a β-lactam ring as part of their chemical structure that can disrupt bacterial cell wall synthesis. Nonetheless, M. catarrhalis produces the enzyme β-lactamase, which can modify the β-lactam ring and thereby renders these antibiotics inactive. Fortunately, the pathogen is still sensitive to most antibiotics used for treating respiratory tract infections. Ciprofloxacin, cefixime, chloramphenicol and amoxicillin-clavulanic acid are examples of antibiotics that are effective against M. catarrhalis infection.
References
Aebi C. 2011. Moraxella catarrhalis – pathogen or commensal? In: Curtis N, Finn A, Pollard AJ, editors. Hot topics in infection and immunity in children vii. New York, NY: Springer New York. p. 107-116. https://link.springer.com/chapter/10.1007%2F978-1-4419-7185-2_9.
Bernhard S, Spaniol V, Aebi C. 2012. Molecular pathogenesis of infections caused by Moraxella catarrhalis in children. Swiss Medical Weekly. [accessed 2021 Nov 13]; 142:w13694. https://pubmed.ncbi.nlm.nih.gov/23136074/. doi:10.4414/smw.2012.13694
Enright MC, McKenzie H. 1997. Moraxella (branhamella) catarrhalis – clinical and molecular aspects of a rediscovered pathogen. Journal of Medical Microbiology. [accessed 2021 Nov 12]; 46(5):360-371. https://www.microbiologyresearch.org/content/journal/jmm/10.1099/00222615-46-5-360;jsessionid=E0PEruR-ZdDu55F0y2vkxuCM.mbslive-10-240-10-26. doi:10.1099/00222615-46-5-360
Goldstein EJC, Murphy TF, Parameswaran GI. 2009. Moraxella catarrhalis, a human respiratory tract pathogen. Clinical Infectious Diseases. [accessed 2021 Nov 12]; 49(1):124-131. https://academic.oup.com/cid/article/49/1/124/371116. doi:10.1086/599375
Murphy TF, Brauer AL, Grant BJB, Sethi S. 2005. Moraxella catarrhalis in chronic obstructive pulmonary disease: Burden of disease and immune response. American Journal of Respiratory and Critical Care Medicine. [accessed 2021 Nov 11]; 172(2):195-199. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2718466/. doi:10.1164/rccm.200412-1747OC
Nordström T, Blom AM, Forsgren A, Riesbeck K. 2004. The Emerging Pathogen Moraxella catarrhalis Interacts with Complement Inhibitor C4b Binding Protein through Ubiquitous Surface Proteins A1 and A2. The Journal of Immunology. [accessed 2021 Nov 12]; 173(7):4598. https://www.jimmunol.org/content/173/7/4598.long. doi:10.4049/jimmunol.173.7.4598
Nordström T, Blom AM, Tan TT, Forsgren A, Riesbeck K. 2005. Ionic Binding of C3 to the Human Pathogen Moraxella catarrhalis is a Unique Mechanism for Combating Innate Immunity. The Journal of Immunology. [accessed 2021 Nov 13];175(6):3628. https://www.jimmunol.org/content/175/6/3628.long. doi: 10.4049/jimmunol.175.6.3628
Su Y-C, Singh B, Riesbeck K. 2012. Moraxella catarrhalis: From interactions with the host immune system to vaccine development. Future Microbiology. [accessed 2021 Nov 11]; 7(9):1073-1100. https://www.proquest.com/docview/1038321799/fulltextPDF/83544917F04C476CPQ/1?accountid=12339. doi:10.2217/fmb.12.80
Verduin CM, Hol C, Fleer A, van Dijk H, van Belkum A. 2002. Moraxella catarrhalis: From emerging to established pathogen. Clinical Microbiology Reviews. [accessed 2021 Nov13]; 15(1):125-144. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC118065/. doi:10.1128/CMR.15.1.125-144.2002
Warnke P, Köller T, Kreikemeyer B, Barrantes I, Mach H, Podbielski A. 2019. Molecular epidemiology study of a nosocomial Moraxella catarrhalis outbreak in a neurological rehabilitation unit. Journal of Hospital Infection. [accessed 2021 Nov 14]; 103(1):27-34. https://www-sciencedirect-com.proxy3.library.mcgill.ca/science/article/pii/S0195670119301884. doi:10.1016/j.jhin.2019.04.019
Yang M, Johnson A, Murphy TF. 2011. Characterization and evaluation of the Moraxella catarrhalis oligopeptide permease as a mucosal vaccine antigen. Infection and Immunity. [accessed 2021 Nov 11]; 79(2):846-857. https://pubmed.ncbi.nlm.nih.gov/21134967/. doi:10.1128/IAI.00314-10
