
by Thomas Donoso and Dennis Park
Introduction
Porphyromonas gingivalis is the opportunistic pathogen largely responsible for periodontitis, an inflammatory oral disease that results in gum damage and tooth loss. The bacterium resides in the oral cavity of humans with hundreds of other bacteria which make up the normal microbiota. This article will explore the pathogenicity of Porphyromonas gingivalis and its several virulence factors.
Disease
Porphyromonas gingivalis is an opportunistic pathogen, meaning it only infects when an imbalance in the normal host microbial flora occurs. Normally, P. gingivalis resides stably with billions of other bacteria in the oral cavity. However, when the ratio or number of the normal microbes is disrupted, it quickly takes advantage of the situation to invade the underlying oral tissue. Inside the oral tissue, P. gingivalis is able to prevent being killed by lysosomes – sacs filled with corrosive and damaging molecules. This gives it enough time to replicate and spread from cell to cell. As the bacteria replicates and spreads, it progressively degrades the nearby tissues using its protein degrading enzymes, one of which is called gingipains, and induces the host’s inflammatory response. This results in the formation of “pockets” around the teeth that is characteristic of periodontitis. As shown in figure 1, a healthy oral cavity has gingiva (gum) that is tightly wrapped around the teeth. However, with periodontitis, the gums are retracted due to proteases and inflammation, therefore resulting in holes and gaps around the teeth. This weakened junction between teeth and gum is what causes the eventual tooth loss during periodontitis.
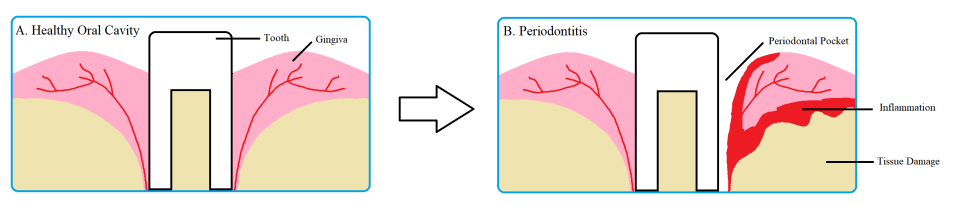
Fig 1. Periodontitis decays gum tissue resulting in periodontal pockets. Healthy teeth are wrapped tightly around by gum tissue. With periodontitis, inflammation and bacterial proteases cause the tissue surrounding the tooth to recede and decay, resulting in “pockets” or gaps. Source: Thomas Donoso and Dennis Park.
Epidemiology
The prevalence of periodontitis is a global phenomenon, with different rates among regions, habits, sex, and race. In 1999, 35% of adults over the age of 30 in the United States were diagnosed with periodontitis. This percentage is correlated to an increase in age. Youth do carry P. gingivalis, however, as studies show that this bacteria was found in cavities with a similar prevalence in all ages. Ultimately, the disease has less effect on younger populations (specifically, less than 30 years old). On top of age, cases of periodontitis have a higher occurrence in those of African, Caribbean, and Mexican descent. In terms of habit, smokers and those who have poorly controlled diabetes are also more susceptible to periodontitis. Lastly, sex is also a contributing factor, as males are more prone to periodontitis than females.
Virulence systems
Porphyromonas gingivalis invades oral tissues using structures called the fimbriae, short hair-like extensions on the surface of the bacterium. The fimbriae attaches to molecules called integrins, specialized receptors on the host tissue, and causes the internal cytoskeleton to be rearranged in the host cell (figure 2A and 2B). The cytoskeleton can be thought of as bones: it supports the cell structure and is involved in cell motility.
Once inside the host cell, the bacterium is enclosed in a phagosome – a sac like compartment (figure 2C). Inside the phagosome, the bacterium cannot survive for long due to two reasons. Firstly, it has limited nutrients to grow, and secondly, the phagosome will soon fuse with a lysosome (another sac filled with corrosive and damaging molecules) and thus, the bacteria will die.
Therefore, P. gingivalis induces the host to undergo autophagy, a highly regulated process that is used to recycle cellular components (figure 2D). During autophagy, the host creates a membrane around cellular components that is to be digested and recycled. The enclosure is later fused with a lysosome, which degrades the components. For reasons not well understood, P. gingivalis can induce the host to undergo autophagy by increasing expression of molecules that regulate this process, such as Beclin-1. The structure that arises from the membrane enclosure around the P. gingivalis (due to autophagy) is called the autophagosome.
Ultimately, inside the autophagosome, the bacteria uses amino acids from the host and replicates using the nutrients (figure 2E). Unfortunately for the bacteria, the autophagosome too will eventually fuse with the lysosome. However, P. gingivalis has cleverly designed methods to delay maturation and fusion of the autophagosome with the lysosome.
The process mentioned above involves various virulence factors, tools used to help induce disease, but one particular virulence factor of interest are the protein degrading enzymes called gingipains produced by P. gingivalis. It can either be membrane bound or secreted.
It is believed that transfer into the autophagosome (figure 2C to 2E) is in part due to the gingipains breaking down the inner membrane of the autophagosome, allowing entry for the bacterium. Also, it is loosely evidenced that gingipains are involved in delaying the fusion between autophagosome and lysosome. For example, experiments by Yamatake et al. 2007 show P. gingivalis mutants that lack gingipains are less observed in autophagosomes and are killed much faster by lysosomes.
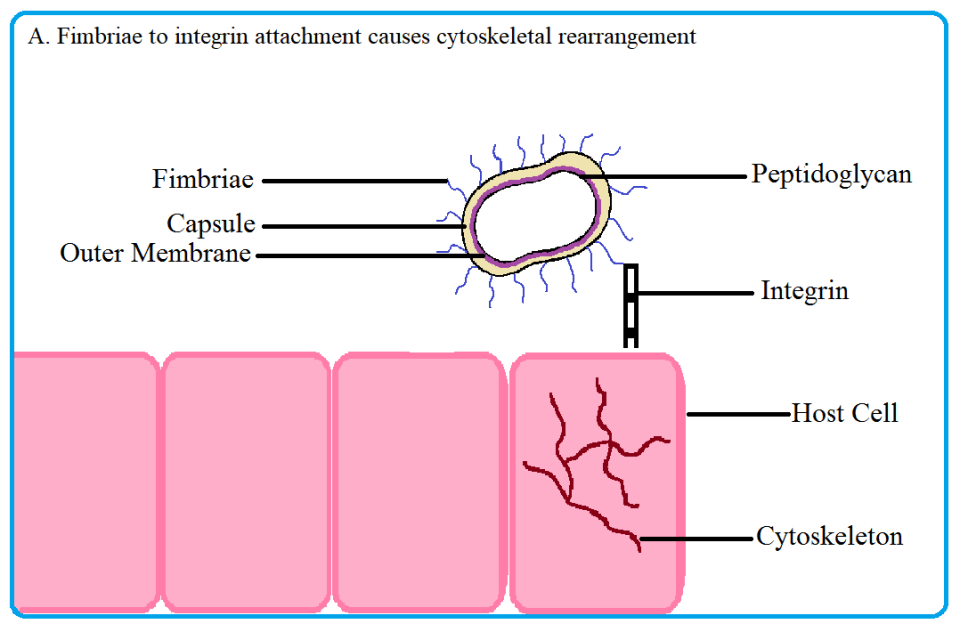
 Fig 2. Porphyromonas gingivalis uses its fimbriae to attach to integrin molecules on the host surface and enter the cell. Upon fimbriae and integrin binding, the cytoskeleton of the host cell rearranges, allowing the pathogen to enter. Once inside, the bacterium induces autophagy in the host to create a nutrient rich niche to replicate in. Source: Thomas Donoso and Dennis Park.
Fig 2. Porphyromonas gingivalis uses its fimbriae to attach to integrin molecules on the host surface and enter the cell. Upon fimbriae and integrin binding, the cytoskeleton of the host cell rearranges, allowing the pathogen to enter. Once inside, the bacterium induces autophagy in the host to create a nutrient rich niche to replicate in. Source: Thomas Donoso and Dennis Park.
Lastly, the most striking fact about gingipains is the wide variety of effects it can have on the host. It is believed that majority of the virulence from P. gingivalis comes from gingipains. Below are some of the effects it can have on a susceptible host and why it contributes to the virulence of P. gingivalis.
| Host molecule | Effect |
| T-Cells* | Cleavage of T-cell receptors prevent proper presentation of antigen (the target of the specific immune system) |
| Pro-inflammatory cytokines** | Pro-inflammatory cytokines are cleaved to weaken the inflammatory response |
| Anti-inflammatory cytokines | Anti-inflammatory cytokines are cleaved to weaken immune regulation by the host |
| Complement*** | Complement molecules are cleaved to prevent recruitment of phagocytes (digestive molecules) and opsonization (marking for digestion) |
| Blood coagulation | Fibrinogen, a protein involved in blood clotting, is degraded to prevent coagulation |
Table 1. Gingipains upset the proper regulation of the host immune response and bodily functions. Gingipains have a wide range of effects within the host. It can prevent proper T-cell function, inflammatory responses and complement activation. Furthermore, it can prevent coagulation to increase migration and replication for the bacterium. * T-Cells are cells of the immune system that help fight off pathogens. It does so either by activating proper responses to deal with the pathogen or by killing cells infected with the pathogen. ** Cytokines are small proteins that act as messengers between cells and tissues. *** Complements are soluble proteins in the blood that can form pores on bacterial membranes or coat bacteria for other immune cells to kill.
Treatment
Before infection, the best method to avoid disease is prevention. Good oral hygiene has been shown to decrease the risk of peritonitis drastically. However, when it is contracted, antibiotics and antiseptics are often prescribed to treat P. gingivalis infection. Positively charged chlorhexidine is an example of an antiseptic used for dental infections, as it kills a broad variety of different bacteria by binding and destroying the negatively charged cell wall. Chlorhexidine is easy to use as it can be administered through a gel or mouthwash, but it may cause irritation.
The alternative, antibiotics, need to be administered through a systemic administration route. This means that the antibiotic needs to be administered through digestion or injection to allow it to reach the deep tissues of the gums. An example of antibiotics used in the treatment are macrolides, such as erythromycin, and ampicillin, which inhibit the synthesis of proteins.
References
Albandar, J. M., Brunelle, J. A., & Kingman, A. (1999). Destructive periodontal disease in adults 30 years of age and older in the United States, 1988-1994. Journal of periodontology, 70(1), 13-29.
Bélanger, M., Rodrigues, P. H., Dunn, Jr, W. A., & Progulske-Fox, A. (2006). Autophagy: a highway for Porphyromonas gingivalis in endothelial cells. Autophagy, 2(3), 165-170.
Bostanci, N., & Belibasakis, G. N. (2012). Porphyromonas gingivalis: an invasive and evasive opportunistic oral pathogen. FEMS microbiology letters, 333(1), 1-9.
Cho, Y. J., Song, H. Y., Ben Amara, H., Choi, B. K., Eunju, R., Cho, Y. A., … & Koo, K. T. (2016). In vivo inhibition of Porphyromonas gingivalis growth and prevention of periodontitis with quorum-sensing inhibitors. Journal of periodontology, 87(9), 1075-1082.
Demmer, R. T., & Papapanou, P. N. (2010). Epidemiologic patterns of chronic and aggressive periodontitis. Periodontology 2000, 53(1), 28-44.
Dorn, B. R., Dunn, W. A., & Progulske‐Fox, A. (2002). Bacterial interactions with the autophagic pathway. Cellular microbiology, 4(1), 1-10.
Gerits, E., Verstraeten, N., & Michiels, J. (2017). New approaches to combat Porphyromonas gingivalis biofilms. Journal of oral microbiology, 9(1), 1300366.
Greene, J. C. (1963). Oral hygiene and periodontal disease. American Journal of Public Health and the Nations Health, 53(6), 913-922.
Griffen, A. L., Becker, M. R., Lyons, S. R., Moeschberger, M. L., & Leys, E. J. (1998). Prevalence of Porphyromonas gingivalis and periodontal health status. Journal of clinical microbiology, 36(11), 3239-3242.
How, K. Y., Song, K. P., & Chan, K. G. (2016). Porphyromonas gingivalis: an overview of periodontopathic pathogen below the gum line. Frontiers in microbiology, 7.
Maezono, H., Noiri, Y., Asahi, Y., Yamaguchi, M., Yamamoto, R., Izutani, N., … & Ebisu, S. (2011). Antibiofilm effects of azithromycin and erythromycin on Porphyromonas gingivalis. Antimicrobial agents and chemotherapy, 55(12), 5887-5892.
McClellan, D. L., Griffen, A. L., & Leys, E. J. (1996). Age and prevalence of Porphyromonas gingivalis in children. Journal of clinical microbiology, 34(8), 2017-2019.
Mysak, J., Podzimek, S., Sommerova, P., Lyuya-Mi, Y., Bartova, J., Janatova, T., … & Duskova, J. (2014). Porphyromonas gingivalis: major periodontopathic pathogen overview. Journal of immunology research, 2014.
Park, M. H., Jeong, S. Y., Na, H. S., & Chung, J. (2017). Porphyromonas gingivalis induces autophagy in THP‐1‐derived macrophages. Molecular oral microbiology, 32(1), 48-59.
Yamatake, K., Maeda, M., Kadowaki, T., Takii, R., Tsukuba, T., Ueno, T., … & Yamamoto, K. (2007). Role for gingipains in Porphyromonas gingivalis traffic to phagolysosomes and survival in human aortic endothelial cells. Infection and immunity, 75(5), 2090-2100.
Yilmaz, Ö., Verbeke, P., Lamont, R. J., & Ojcius, D. M. (2006). Intercellular spreading of Porphyromonas gingivalis infection in primary gingival epithelial cells. Infection and immunity, 74(1), 703-710.

This is so well done, detailed and in depth. May we all take heed.
I am very proud of you Thomas.
Grandma and Grandpa
LikeLike